Общество
«Знание» РСФСР
Ленинградская
организация
Доктор
биологических наук
Н.
В. Голиков

МЕХАНИЗМЫ
РАБОТЫ МОЗГА
ЛЕНИНГРАД
1968
5А2.991.7
Г-604
Голиков Н. В.
Г-604
Механизмы работы мозга
Л.,
«Знание», 1968. 3 л.
(О-во
«Знание» РСФСР. Ленинградская организация)
10250
экз. 8 коп. Перед загл: авт.:
Н.
В. ГОЛИКОВ, проф.
Крупный учёный, представитель
ленинградской университетской
физиологической
школы профессор Н.В. Голиков обобщает
главнейшие
итоги и раскрывает перспективы развития
нейрофизиологических
исследований механизмов деятельности
нервной
системы.
Автор подчёркивает теснейшую
связь физиологического
учения
Введенского и Ухтомского о механизмах нервной
деятельности
с задачами и перспективами современной
нейрофизиологии.
Брошюра рассчитана на читателей,
знакомых с основами
физиологии
нервной системы; особый интерес представляет
для
биологов, физиологов и врачей различных специальностей,
а
также студентов медицинских, педагогических и биологических
факультетов.
5-3-12
32-БЗ-19-68 5А2.991.7
Николай
Васильевич Голиков
МЕХАНИЗМЫ
РАБОТЫ МОЗГА
(Бланк
заказов, № 19, 1968 г.)
Научный
редактор канд. медицинских наук Н. Н. Василевский
Редактор
издательства В. В. Николаева
Обложка
работы В, А. Политова
Технический
редактор М. Я. Петрова
Корректор
Б. Р. Трахтман
М-38907.
Сдано в набор 22/Ш-68 г. Подписано к печати 20/VI1I-68 г.
Формат
84X108V32- Печ. л. 3,0 (Усл.-печ. л. 2,5). Уч.-изд. л. 2,5,
Бумага
типографская № 3. Тир. 10 250 экз. Зак. №505. Цена 8 коп.
Ленинградская
организация
общества
«Знание» РСФСР, Ленинград, Д-104, Литейный пр., 42.
Типография
№ 4 Управления по печати Ленгорисполкома, г. Пушкин
В процессе
эволюции многоклеточных организмов происходила постепенная дифференциация
клеток, возникали специализированные ткани и органы, обеспечивавшие более
совершенное и более экономное осуществление жизненных функций.
В каждой клетке
под влиянием раздражения наступают фазно протекающие изменения тонкой
структуры, проницаемости клеточных мембран, степени дисперсности биополимеров,
электропроводности, электрического потенциала, интенсивности обмена веществ,
чувствительности к раздражениям, скорости протекания реакций и т. д. В
специализированных клетках при их возбуждении на передний план выступают либо
структурные изменения, обусловливающие возникновение сокращения (мышечныё
клетки), либо химические изменения — изменения обмена веществ, обусловливающие
образование и выделение секрета (секреторные клетки), либо перемещение ионов,
связанные с изменением электрического потенциала и возникновением разряда
(электрический орган электрических рыб) и т. д.
Таким образом,
компоненты общебиологической реакции, возникающей в каждой клетке в ответ на
раздражение, оказываются наиболее резко выраженными в специализированных
клетках. При этом другие компоненты реакции подавляются, как бы отступая на
задний план.
Некоторые
клеточные элементы наружного покрова тела (эктодермы) уже у примитивных
многоклеточных животных приобрели высокую чувствительность и развили
способность проводить вспышки возбуждения без затухания (декремента, т.е. без
изменения амплитуды импульса), по своим отросткам к другим клеткам и
регулировать их деятельность. Специализированные высокочувствительные клетки с
длинными отростками, первыми реагирующие на раздражающие воздействия не только
местной реакцией, как другие клетки, но
и распространяющимися по этим отросткам импульсами возбуждения и длительно
сохраняющие специфические следы раздражений, получили название нервных клеток,
или нейронов.

Рис.
1. Типы нейронов.
A — пирамидальная клетка Беца (из
коры мозга), B — клетка Пуркинье (из коры
мозжечка), C — униполярная клетка
(псевдоуниполярная), D — мультиполярная клетка (из
спинного мозга), E — ганглиозная клетка.
Весьма
своеобразную эволюцию претерпели окончания отростков нервных клеток: одни
окончания избирательно повысили свою чувствительность к определённым физическим
или химическим раздражениям, стали рецепторными, воспринимающими, другие
окончания (аксонов) столь же значительно развили свою способность
воздействовать на другие клетки. Этот тип окончаний образует на иннервируемых
ими клетках специальные образования — синапсы.
Совокупность
нейронов, связанных своими отростками друг с другом, с рецепторами (органами
чувств) и исполнительными органами (мышцами, железами), получила название
нервной системы.
Возникновение
нервной системы и дальнейшая специализация нейронов и их рецепторных и
эффекторных окончаний обеспечили появление новых приспособительных реакций и
быстрое развитие животных. Нервная система первой отвечает на раздражающие
воздействия среды, регулирует состояние всех тканей и органов и объединяет эти
ткани и органы в целостный организм, обладающий более высокой чувствительностью
и большей скоростью реакций по сравнению с организмами, не обладающими нервной
системой. По мере развития животных нервная система всё более
совершенствовалась, обеспечивала быстроту и точность всё усложняющихся реакций
организма и всё большую слаженность (интеграцию) работы его органов.
У низших
многоклеточных организмов обнаружены лишь отдельные нервные (чувствительные)
клетки с ещё слабо развитой рецепторной и эффекторной функцией (губки). Далее у
кишечнополостных уже образовалась сеть нейронов, распределённая по всему телу
животного, элементы которой связаны своими отростками друг с другом, с
рецепторами и исполнительными органами (так называемый диффузный тип строения
нервной системы). В нервной системе моллюсков нейроны сконцентрировались в узлы
(церебральный, висцеральный и педальный ганглии), образуя ганглиозную нервную
систему. У червей и членистоногих нервные ганглии составляют уже парную цепочку
или лестницу, причём в каждом сегменте (метамере) тела находится один парный
ганглий, связанный со своими рецепторными и зффекторными аппаратами и с
ганглиями других сегментов (лестничный тип строения нервной системы). Ганглии
переднего конца тела (надглоточный и подглоточиый узлы), первыми воспринимающие
раздражающие воздействия, получают преимущественное развитие и подчиняют себе
нижележащие сегменты нервной системы, У позвоночных животных (рыбы, амфибии,
рептилии и млекопитающие) количество нервных клеток значительно возрастает,
сегментарно расположенные парные ганглии сливаются и образуют нервную трубку
(трубчатый тип строения нервной системы). Из переднего конца первичной нервной
трубки, представленного тремя мозговыми пузырями, развиваются передний, средний
и задний отделы мозга, расположенные в полости черепа, а из заднего конца этой
трубки формируется спинной мозг.
Аксоны (длинные
отростки) клеток нервной системы имеют большое количество коллатералей (боковых
ветвей), посредством которых каждая нервная клетка может связываться с тысячами
других, чем и обеспечивается широкая возможность межнейронных взаимоотношений.
Каждый метамер
тела связан не только со своим сегментом нервной системы, но и с выше и ниже
лежащими (принцип перекрытия, обеспечивающий надёжность нервной регуляции и
связи).
Количество
афферентных (чувствительных) нервных клеток значительно больше количества
эфферентных (двигательных, оконечных) клеток. К каждому вставочному и
эфферентному нейрону подходят аксоны от многих чувствительных и других
вставочных нейронов (принцип конвергенции или принцип общего пути в строении
нервной системы). Эта особенность в строении нервной системы имеет важное
значение для координации её рефлекторной деятельности. В пунктах конвергенции
(схождения) путей происходит взаимодействие нервных импульсов, приходящих из
разных рецепторов и нейронов, в результате чего изменяется, регулируется
реакция нейронов, усиливается или подавляется их деятельность, развиваются
процессы возбуждения или торможения.
Реакции нервной
системы на раздражающие воздействия называются рефлексами, актами отражения,
Каждый рефлекс осуществляется более или менее сложной цепочкой нейронов,
получившей название рефлекторной дуги. Простейшая рефлекторная дуга состоит
всего из двух нейронов — афферентного (чувствительного) и эфферентного
(двигательного). Более сложные рефлекторные дуги состоят из значительного
количества нейронов, при этом между афферентным и эфферентным нейронами имеются
вставочные или промежуточные нейроны. Такие рефлекторные дуги с многозвенными
синаптическими связями получили название полисинаптических. В этих дугах
вставочные и эфферентные нейроны являются местом взаимодействия конвергирующих
к ним импульсов возбуждения.
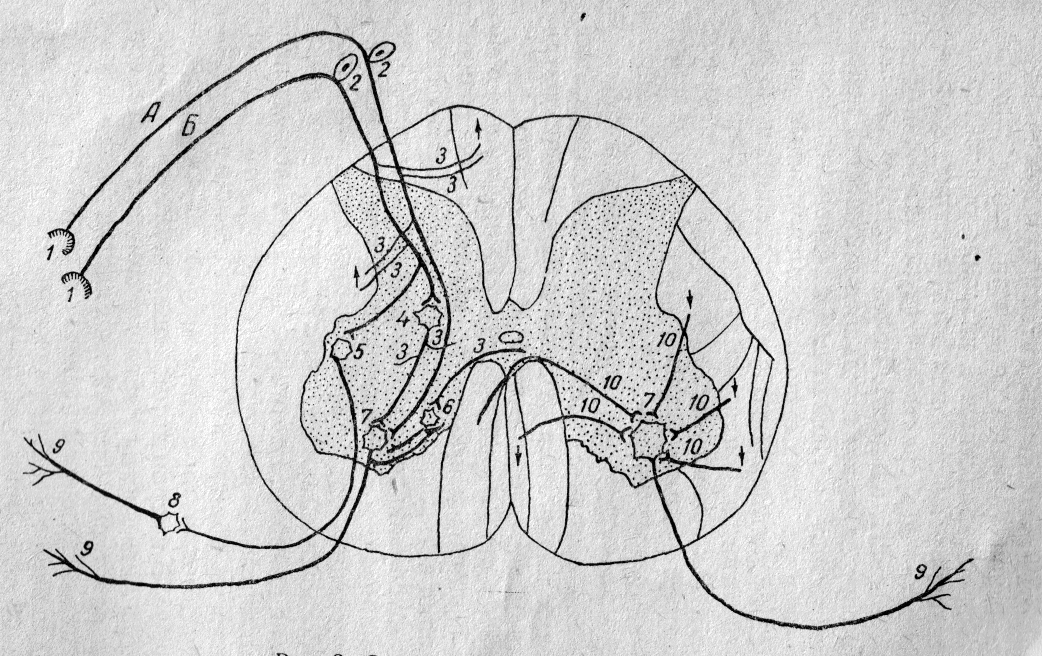
Рис.
2. Схема межнейронных связей.
А — моносинаптическая
рефлекторная дуга, Б — полисинаптическая рефлекторная дуга. 1 — рецептор, 2 —
чувствительный нейрон, 3 — коллатерали, 4 — вставочный нейрон, 5 — вставочный
вегетативный нейрон, 6 — клетка Реншоу, 7 — двигательный нейрон, 8 — эфферентный
вегетативный нейрон, 9 — эффектор, 10 — супрасегментарные связи.
Состояние
возбуждения нейрона, выражающееся в его импульсных разрядах, и состояние
торможения, проявляющееся в подавлении этих разрядов, возникают в зависимости
от характера импульсации (частоты, длительности, степени синхронизации),
специализации нейронов, их синапсов и исходного уровня их состояния.
Специализация вставочных нейронов и их синаптических окончаний обусловливает
тот факт, что эти нейроны при широком диапазоне изменений частоты и
продолжительности импульсации оказывают обычно лишь однозначное — либо
возбуждающее, либо тормозящее — действие на другие нейроны. Некоторые типы
вставочных нейронов специализировались таким образом, что их окончания
(синапсы) при своей активации обычно лишь снижают возбудимость, тормозят
нейроны, на которые они воздействуют. Примером тормозящих вставочных нейронов
могут служить клетки Реншоу в спинном мозге, корзинчатые и некоторые звёздчатые
клетки в головном мозге. Специализация обеспечивает надёжность возникновения
однозначного специфического влияния активации этих нейронов. Лишь при
чрезмерных раздражениях и значительных отклонениях функционального состояния
нервных клеток от нормального уровня наблюдается извращение обычного
специфического эффекта стимуляции в противоположный.
При изучении
механизма влияния нейронов друг на друга и на другие специализированные клетки
тела необходимо учитывать, что каждая клетка влияет на другую не иначе, как
раздражая её электрическими (петля местного потенциала) или
химическими (медиаторы, метаболиты) компонентами своего возбуждения. В
связи с этим для понимания механизмов нервной деятельности особое значение
имеют закономерности раздражения, лежащие в основе межцентральных и
регулирующих влияний.
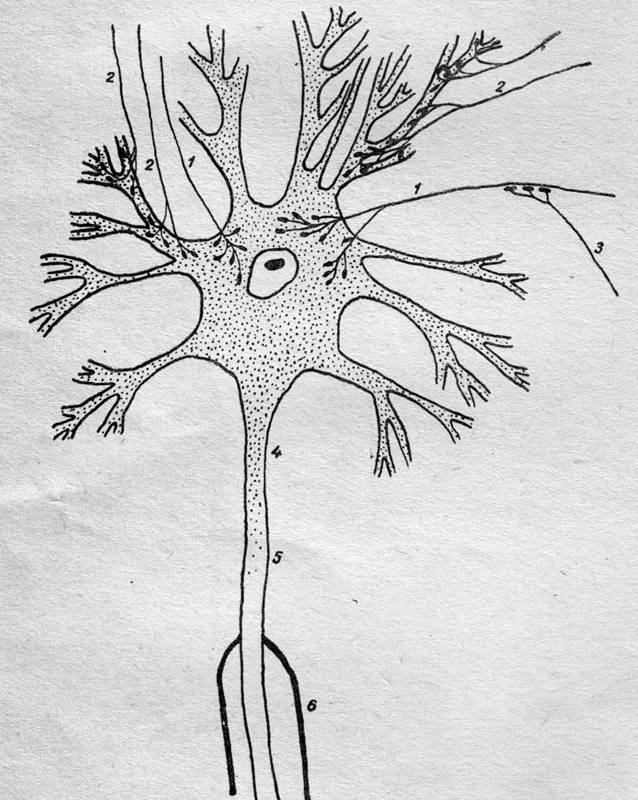
Рис.
3. Схема синаптических окончаний.
1 — аксо-соматические синапсы, 2
— аксо-дендритные синапсы, 3 — аксо-аксонные синапсы, 4 — воронка аксона и
аксон, 5 — немиелизированная часть аксона, 6 — миелизированная часть аксона.
В этом плане
особенно важное значение имеют открытия и обобщения основателей университетской
физиологической школы Ленинграда — Н.Е. Введенского и А.А. Ухтомского, которые
внесли существенный вклад в развитие современной нейрофизиологии. Изучая
процессы возбуждения и торможения на нервно-мышечном препарате и в относительно
простых рефлекторных дугах, Н.Е. Введенский показал, что для возникновения
возбуждений необходима определённая, оптимальная величина раздражения. Слишком
сильные, пессимальные, раздражения снижают возбудимость, прекращают проведение
как в нервно-мышечном аппарате, так и в рефлекторных дугах (закон оптимума
величины раздражения, необходимой для вызова возбуждения). Телефоническое
исследование феноменов оптимального (возбуждающего) и пессимального
(тормозящего) действия раздражений позволило Н.Е. Введенскому показать, что при
возникновении торможения под влиянием пессимального раздражения электрические
компоненты элементарных вспышек возбуждения начинают протекать все медленнее,
эти вспышки перестают распространяться по нервным и мышечным волокнам.
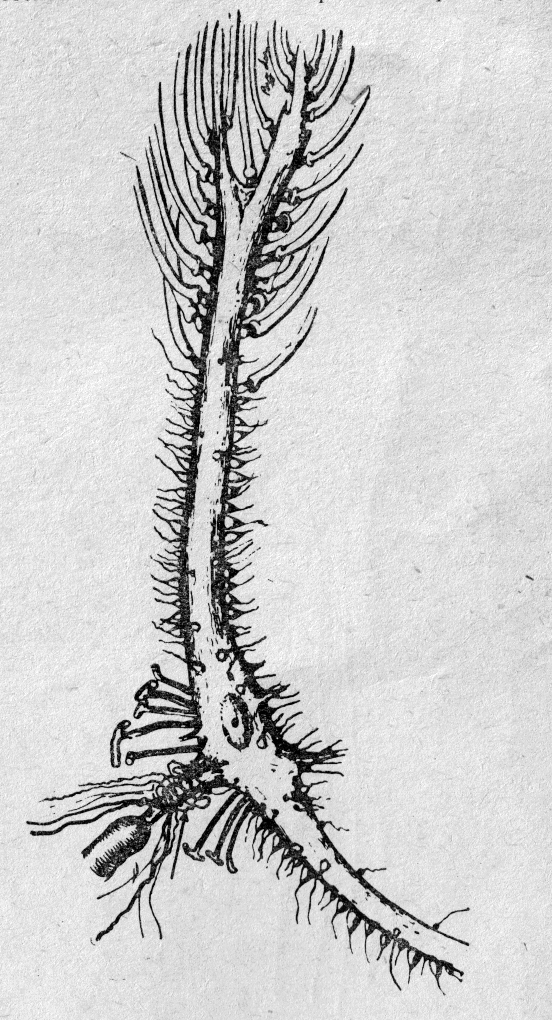
Рис.
3a. Синапсы на нервной клетке.
Показав
соответствие электрических и функциональных изменений в клетках и волокнах,
Н.Е. Введенский обозначил скорость протекания элементарных вспышек возбуждения
термином «функциональная подвижность», или «лабильность», и предложил измерять
её максимальным числом электрических компонентов вспышек возбуждения,
возникающих в течение одной секунды в точном соответствии с ритмом
максимального раздражения. Удобной мерой лабильности является также
продолжительность одной элементарной вспышки возбуждения, соответствующая
длительности тока действия (спайка).
Прекращение
проведения импульсов возбуждения (торможение) Введенский связывал со снижением
лабильности и наличием стойкого местного, нераспространяющегося возбуждения в
зоне раздражения.
Современные
электрофизиологические исследования показывают, что более древние по
происхождению местные возбуждения могут существовать и даже быть более
выраженными при утрате способности к проведению импульсов возбуждения.
Прекращение проведения (торможение системной реакции) может сочетаться с
наличием местных возбуждений клеток и волокон. Так, медленные колебания
электрического потенциала в различных отделах мозга, соответствующих местным приступам
возбуждения, могут регистрироваться после утраты рефлекторной возбудимости и
исчезновения быстрой электрической активности, связанной с возникновением
импульсов возбуждения, распространяющихся по аксонам.
Однако еще при
жизни Н.Е. Введенского были известны факты, показывавшие наличие снижения
возбудимости и прекращения проведения при повышении лабильности, например, при
возникновении анэлектротона, когда на ткань воздействует анод постоянного тока.
Известны были также факты снижения рефлекторной возбудимости под влиянием
слабых раздражений. Так, И.П. Павлов показывал, что для выработки условного
рефлекса необходима определённая величина условных и безусловных раздражений.
Не только более сильные раздражения, но и раздражения более слабые, чем оптимальные
для выработки рефлекса, ведут к снижению возбудимости и возникновению
тормозного процесса.
Эти факты, а
также описанные И.М. Сеченовым различные формы торможения побудили А.А.
Ухтомского (1911), Л.Л. Васильева (1924), Э.Т. Брюкке (1922) и других различать
две формы прекращения проведения импульсов возбуждения (возникновения
торможения) — первичное торможение, соответствующее анэлектротоническому
снижению возбудимости, и вторичное торможение, соответствующее катодической
депрессии, состоянию пессимума и парабиоза Н.Е. Введенского.
А.А. Ухтомский и
его сотрудники дополнили учение Введенского о возбуждении и торможении, показав
наличие начального повышения и последующего снижения физиологической
лабильности клеток и волокон под влиянием раздражений. В действии раздражителей
на клетки и волокна были обнаружены двуфазные изменения лабильности и
трёхфазные изменения возбудимости (Голиков, 1933). Первичное, непосредственное
торможение оказалось идентичным первой фазе неспецифической реакции клеток и
волокон на длительные раздражающие воздействия, особенно ярко выступающей при
слабых раздражениях.
Используя схему
изменений нормальной поляризации клеточных мембран при возникновении
возбуждения, предложенную в начале нынешнего века Бернштейном (1912), и
рассматривая нормальную поляризацию клеток как степень их ионной асимметрии с
окружающей средой, можно сопоставить уровень местного возбуждения Н.Е.
Введенского и уровень поляризации раздражаемых клеток и волокон.
Была показана
связь между величиной поляризации (потенциалом покоя) и уровнями возбудимости и
лабильности клеток и волокон. Лабильность, скорость клеточных элементарных
реакций, изменялась параллельно с изменением уровня поляризации, повышаясь при
возрастании потенциала покоя, при гиперполяризации клетки, и снижаясь при
уменьшении потенциала покоя, при деполяризации клетки или волокна.
Уровень
возбудимости клеток и волокон оказался наиболее высоким при определённой
средней величине потенциала покоя, обычно соответствующей умеренной
деполяризации. Как повышение, так и чрезмерное снижение поляризации от
оптимального его уровня ведет к снижению и исчезновению возбудимости,
способности к проведению. Для наличия проведения и возбудимости необходима
определённая величина поляризации и лабильности клеток и волокон (Голиков,
1933). Как гиперполяризация, так и глубокая деполяризация снижают и уничтожают
возбудимость и проведение.
Эта
закономерность связана с открытым Н.Е. Введенским законом оптимальной величины
раздражения, необходимой для возникновения возбуждения, и демонстрирует наличие
специфического для живых образований начального трофического действия
раздражителей, сменяющегося последующим возбуждающим, а затем тормозящим их
действием.
Использование
методики внутриклеточного отведения потенциалов подтвердило закономерную связь
уровня возбудимости и способности к проведению с величиной внутриклеточного
потенциала. Как в нейронах, так и в нервных и мышечных волокнах значительное
увеличение внутриклеточного потенциала (гиперполяризация), а также значительное
снижение этого потенциала (глубокая деполяризация); исключают возможность
возникновения распространяющихся импульсов возбуждения. Средняя величина
поляризации (умеренная деполяризация), напротив, связана с повышением
возбудимости и облегчением проведения.
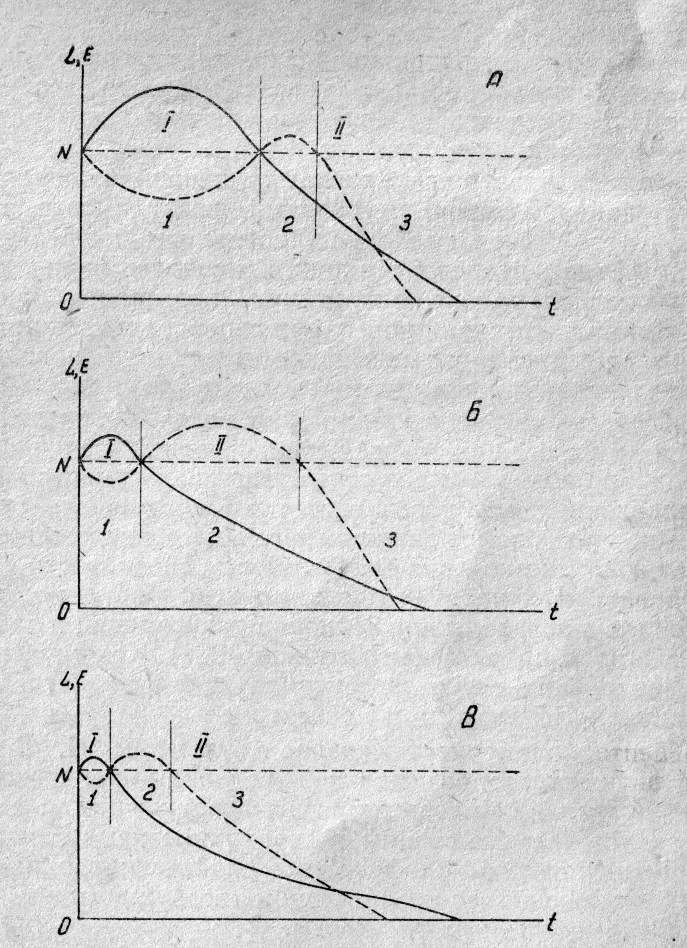
Рис.
4. Схема изменения поляризации, лабильности и возбудимости нервной ткани при
длительных раздражениях.
По оси абсцисс — время действия
раздражителя, по оси ординат — изменения поляризации и лабильности (сплошная
линия) и изменения возбудимости (прерывистая линия). I — фаза повышения поляризации и
лабильности, II — фаза снижения поляризации и
лабильности. 1 — фаза первичного снижения возбудимости, 2 — фаза повышения
возбудимости, 3 — фаза вторичного снижения возбудимости. А,Б, В — действие
факторов, подчёркивающих и затягивающих ту или иную фазу неспецифической
реакции.
В условиях этой
методики подтверждён также и тот факт, что при длительных раздражениях в
клетках и волокнах возникают двуфазные изменения лабильности и поляризации
(повышение — снижение) и обычно трёхфазные изменения возбудимости (снижение —
повышение — снижение). Трёхфазные изменения возбудимости соответствуют
возникновению первичного адаптационного гиперполяризационного торможения
(анэлектротонический синдром) последующему возбуждению (катэлектротонический
синдром), переходящему во вторичное деполяризационное, запредельное торможение
(синдром катодической депрессии).
В низколабильных
структурах при тех же длительных раздражениях возбудимость изменяется, так же
как поляризация и лабильность, двуфазно (повышение — снижение). Начальное
положительно трофическое действие сказывается здесь не в снижении, а в
повышении возбудимости, но связанном с тем же повышением лабильности и
поляризации.
Большое
количество новых фактов показывает, что специфика раздражителей и специализация
раздражаемых структур могут подчёркивать и затягивать ту или иную фазу общей
(неспецифической) реакции клеток и волокон на раздражающие воздействия.
Специализация рецепторов, клеток и синапсов расширяет диапазон специфического
(однозначного) действия одних раздражителей и ограничивает возможность
эффективного действия других «неадекватных» раздражителей.
Чувствительность
специализированных структур к определённому раздражителю может чрезвычайно
возрастать по сравнению с чувствительностью других, неспециализированных
структур к тому же раздражителю. Так, дозы медиатора, совершенно неэффективные
для других клеток и свободных от синапсов зон данной клетки, оказываются резко
возбуждающими для субсинаптических рецептивных структур. На одну и ту же порцию
медиатора, выделяемую нервными окончаниями в синаптическую щель, одни клетки
отвечают увеличением внутриклеточного потенциала (гиперполяризацией) и
снижением возбудимости, возникновением торможения, а другие клетки, напротив,
реагируют уменьшением внутриклеточного потенциала (умеренной деполяризацией), повышением
возбудимости, возникновением импульсного разряда, распространяющегося по
аксону.
Каждый тип
специализированных синаптических аппаратов может вызывать либо гиперполяризацию
и первичное торможение (тормозные синапсы), либо умеренную деполяризацию и
возбуждение (возбуждающие синапсы), либо сразу глубокую деполяризацию и
пессимальное, запредельное торможение (аксо-аксонные, резко деполяризующие,
тормозные синапсы). Однако в определённых условиях обычная специфическая
реакция может изменяться с выявлением общих закономерностей фазного развития
неспецифической клеточной реакции.
Реакция нейронов
зависит не только от специфики синаптических аппаратов, но и от частоты, силы и
длительности импульсации, степени её синхронизации и от уровня функционального
состояния реагирующих клеток.
При изменении
величины стимуляции и её особенностей или при изменении состояния реагирующих
нейронов или других клеток иннервируемого органа обычный эффект стимуляции
может изменяться или извращаться. Так, у ненаркотизированного животного слабое
раздражение центрального конца блуждающего нерва вызывает первичное торможение
дыхательного центра. Такое же раздражение того же нерва у наркотизированного
животного влечет за собой длительное возбуждение дыхательного центра. Более
сильные раздражения у ненаркотизированного животного вызывает возбуждение
дыхательного центра. Те же раздражения у наркотизированного животного вызывают
вторичное, запредельное торможение этого центра.
В соответствии с
законом Введенского, по которому явления пессимума (функционального парабиоза,
вторичного торможения) более легко возникают при снижении уровня лабильности
клеток и волокон, деполяризационное торможение легче возникает в малолабильных,
значительно деполяризационных клетках и волокнах. Напротив, первичное
гиперполяризационное торможение и явления адаптации нарушаются и исчезают при
снижении лабильности и поляризации соответствующих клеток и волокон. В первые
дни постнатального развития котят перекрестное первичное гиперполяризационное
торможение в двунейронных рефлекторных дугах спинного мозга не возникает;
вместо торможения возникает эффект возбуждения, и лишь в последующие дни
появляется тормозной эффект наряду с повышением уровня лабильности и
поляризации двигательных нейронов. Нормальная остановка сердца при раздражении
блуждающего нерва заменяется усиливающим и учащающим сердечную деятельность
эффектом этого раздражения на ослабленном или отмирающем сердце.
Возрастание
физиологической лабильности в процессе фило- и онтогенетического развития (А.А.
Ухтомский, 1937) в настоящее время многократно подтверждено. Имеются данные,
показывающие увеличение уровня поляризации нейронов в процессе
онтогенетического развития (В.С. Шевелева, 1965). Появление и возрастание
возбудимости по мере созревания нейронов симпатического ганглия сменяется
вторичным её снижением при достижении высокого уровня поляризации. Эти данные
соответствуют наличию закономерных соотношений между уровнями поляризации,
лабильности и возбудимости нервных клеток и волокон.
Специфические
гиперполяризующие факторы снижают возбудимость и вызывают первичное
гиперполяризационное торможение только в структурах с высоким уровнем
лабильности и поляризации. В структурах с низким уровнем лабильности и
поляризации те же факторы вызывают повышение возбудимости и возникновение
возбуждения. Специфические деполяризующие факторы вызывают повышение
возбудимости и возбуждение только в структурах с высоким уровнем лабильности и
поляризации. В структурах с низким уровнем лабильности и поляризации эти
факторы вызывают снижение возбудимости и возникновение деполяризационного
торможения.
Как центральные,
так и периферические эффекты действия нервных импульсов обусловлены не только
специализацией синапсов и рецептивных структур, но и величиной импульсации
(частотой и количеством нервных импульсов и степенью синхронизации их
действия), а также уровнем функционального состояния реагирующих клеток.
Реакции нервной системы на раздражающие воздействия подчиняются общим
закономерностям раздражения. Импульсы возбуждения, возникающие в результате
раздражения рецепторов, нервных волокон или нервных клеток, сами являются
раздражителями как для соседних участков
клетки или волокна, так и для других клеток. Проведение импульса
возбуждения по нервному волокну, как это было показано Германом в конце
прошлого века и неоднократно подтверждено современными электрофизиологическими
исследованиями, является результатом последовательного электрического
раздражения соседних звеньев этого волокна электрическими потенциалами (токами
действия), возникающими при появлении вспышек возбуждения в предыдущем звене
(Тасаки). Нервный импульс, приходящий в синаптическое окончание аксона,
раздражая его током действия, вызывает в нем вспышку местного возбуждения,
сопровождающуюся возникновением электрического потенциала и выделением
метаболитов (медиатора) в синаптическую щель в зоне контакта этого окончания со
следующей клеткой или её отростками и обусловливает химическое или
электрическое раздражение последней.
Зависимость
рефлекторных реакций от величины раздражений в особенности отмечалась И.М.
Сеченовым и Н.Е. Введенским, показавшими, что сильные раздражения вместо
возбуждения могут вызывать возникновение состояния торможения. H. Бубнов и Р.
Гейденгайн (1881), Ч. Шеррингтон (1909), И.С. Беритов (1916), И.П. Павлов
(1924) демонстрировали тот факт, что и слабые раздражения вместо ожидаемого
повышения возбудимости и возбуждения могут вызывать снижение возбудимости и
возникновение тормозных состояний.
Даже при
отсутствии нарочитых раздражений рецепторы отправляют в центральную нервную
систему редкие асинхронные импульсы возбуждения (фоновая афферентная
импульсация). Нервные клетки также периодически отправляют редкие асинхронные
импульсы возбуждения к другим нервным клеткам и в исполнительные органы
(фоновая эфферентная, или субординационная импульсация, или фоновая
межцентральная импульсация). Эта фоновая импульсация обычно снижает
возбудимость и повышает поляризацию и лабильность подвергающихся её действию
клеток. Ч. Шеррингтон показал, что перерезка задних корешков и выключение
притока афферентной импульсации в соответствующие сегменты спинного мозга
вызывают повышение возбудимости двигательных нейронов спинномозговых
рефлекторных дуг деафферентированных сегментов (1909). К. Кунстман и Л.А.
Орбели (1924) подтвердили этот факт и показали, что после перерезки задних
корешков деафферентированная конечность начинает отвечать рефлекторными
сокращениями мышц на каждый залп импульсов, иррадиирующих из дыхательного
центра. Аналогичное повышение возбудимости мотонейронов спинномозговых
рефлекторных дуг было показано при децеребрации И.С. Беритовым (1916). Лишённые
фоновой субординационной импульсации из переднего и среднего мозга, мотонейроны
резко повышали свою возбудимость и отвечали усилением тонических асинхронных
импульсных разрядов при каждом вдохе.
Снижающее
возбудимость и повышающее лабильность мышечных волокон влияние субординационной
асинхронной импульсации было подчеркнуто Монье и Джаспером (1934), показавшими,
что после денервации наступает повышение возбудимости, удлинение хронаксии и
рефрактерной фазы мышечных волокон. Субординирующее влияние сравнивалось ими с
влиянием анэлектротона. Большой материал, показывающий снижающее возбудимость
действие субординационной импульсации, приведен В. Кенноном и А. Розенблютом в
их монографии «Повышение чувствительности денервированных структур» (1951).
Нужно заметить,
однако, что длительное устранение привычной фоновой импульсации ведет к
вторичному снижению возбудимости и развитию глубокого деполяризационного
торможения, несмотря на отсутствие чрезмерных раздражений. Ещё И.М. Сеченов
описал возникновение глубокого сонного состояния при резком уменьшении притока
раздражений из окружающей среды. Больной с обширными поражениями органов чувств
почти круглые сутки спал, пробуждаясь лишь на время еды и отправления
естественных потребностей. В лаборатории И.П. Павлова К.С. Абуладзе и другими
было показано, что после одномоментной перерезки зрительных, слуховых и
обонятельных нервов у собак развивалось глубокое тормозное состояние.
Всё это
показывает, что постоянные слабые раздражающие воздействия оказывают
трофическое влияние, повышающее лабильность и поляризацию нервных клеток.
Выключение раздражающих влияний ведёт к прогрессирующему снижению лабильности и
поляризации, к начальному повышению и последующему снижению возбудимости и,
наконец, к развитию глубокого деполяризационного торможения и даже к смерти.
Состояние нервной системы регулируется раздражающими воздействиями, постоянный
приток раздражений необходим для нормальной нервной деятельности. Как
недостаточный приток раздражений, так и чрезмерные раздражения ведут к снижению
и исчезновению возбудимости и развитию деполяризационного торможения, иногда
предваряемого возникновением этапа повышенной возбудимости и возбуждения.
Отмечая важное
значение межцентральных влияний, А.А. Ухтомский уже в начале нашего века
показал, что повышение возбудимости отдельных нервных центров влияет на
состояние других центров и может существенно изменить, обычные рефлекторные
реакции нервной системы. Реакция нервного центра на раздражающие воздействия
определяется не только величиной и спецификой раздражения и не только уровнем
функционального состояния раздражаемого центра, но и уровнем функционального
состояния других центров.
Изучая
зависимость эффектов раздражения коры от побочных центральных влияний, А.А.
Ухтомский обнаружил, что на иррадирующие в нервной системе импульсы возбуждения
первыми отвечают группы нейронов, находящиеся в состоянии повышенной возбудимости.
Эти нейроны отвечают приступом возбуждения даже на раздражения, адресованные
другим нервным центрам. В это время другие нервные центры оказываются
заторможенными, теряющими способность отвечать импульсным разрядам даже на
раздражения, непосредственно на них действующие. Готовые к импульсному разряду
очаги местного возбуждения, возникшие в результате предыдущих раздражений,
предопределяют характер рефлекторных реакций на последующие раздражения и
обусловливают наличие тех или иных тенденций, готовности нервной системы к
направленным реакциям. Межнейронные взаимодействия могут существенным образом
изменять характер рефлекторных реакций нервной системы на текущие раздражения.
Открытые А.А.
Ухтомским закономерности иррадиации импульсов возбуждения в нервной системе и
закономерности межцентральных
взаимоотношений были им обобщены в виде основного принципа нервной
деятельности — принципа доминанты в работе нервных центров. В этом обобщении
собственные экспериментальные данные были им сопоставлены с огромным
литературным материалом современной нейрофизиологии и электрофизиологии.
Выдвигая принцип
доминанты, А.А. Ухтомский широко использовал учение Н.Е. Введенского о местном
возбуждении, принцип общего пути в строении нервной системы, выдвинутый
известным английским физиологом Ч. Шеррингтоном, и представления И.М. Сеченова
о роли следов раздражений в нервной деятельности.
По А.А.
Ухтомскому, каждый рефлекторный акт осуществляется определённой структурой,
констелляцией нервных центров, часто расположенных в различных этажах нервной
системы, но обладающих единством действия. Длительное возбуждение придает
функциональной констелляции нервных центров главенствующее, доминирующее
значение в поведении человека и животных. Центры, входящие в состав
доминирующей констелляции, характеризуются повышенной возбудимостью, типичной
для наличия местного возбуждения, повышенной способностью к суммации,
стойкостью, инерционностью состояния возбуждения и оказывают сопряжённо
тормозящее действие на другие центры, не вошедшие в состав данной констелляции.
Единство
действия доминирующей констелляции центров, по мысли Ухтомского, связано с
единым ритмом и темпом активности центров — участников этой констелляции. Одни
и те же отдельные центры или группы нейронов могут входить в состав различных
доминирующих констелляций, причем вхождение в данную констелляцию или
выключение из неё тех или иных отдельных центров определяется способностью этих
центров усваивать единый темп и ритм активности всей доминирующей системы
совозбуждённых центров. Формирование доминирующих констелляций нервных центров
связано с достаточно длительным действием оптимальных, рефлекторных и
гуморальных раздражений.
Угашение,
торможение доминантной установки происходит либо в результате устранения
раздражителей, вызвавших и поддерживающих состояние доминанты (её угасание),
либо в результате чрезмерного усиления этих раздражителей (переподкрепление,
переход центров из состояния доминирующего возбуждения в состояние
запредельного торможения, парабиоза), либо в порядке сопряжённого торможения
при возникновении и подкреплении другой доминантной констелляции, несовместимой
с первой.
Следы пережитых
доминант длительно сохраняются в высших отделах нервной системы, и при
определённых условиях каждая доминантная установка может вспыхивать вновь,
репродуцироваться — либо эскизно (в виде воспоминания), либо полностью.
В каждый данный
момент в центральной нервной системе может существовать лишь одна активная
доминантная констелляция центров (нейронных групп), относительно простая или
весьма сложная, способная осуществлять ряд одновременных и последовательных
актов определённой направленности. В единую доминантную констелляцию могут
вовлекаться дополнительные группы нейронов, деятельность которых способна
синхронизироваться с ритмом и темпом деятельности ведущих центров.
Одновременное
осуществление ряда актов иногда неправильно рассматривается как сосуществование
двух или нескольких активных доминантных реакций. На самом деле, как показывают
данные физиологии труда и экспериментальной психологии, в этих случаях имеет
место образование более сложной системы взаимно связанных реакций,
осуществляемых более сложной, но единой доминирующей констелляцией нервных
центров. Артист, например, при исполнении роли может одновременно осуществлять ряд
сложных действий, однако все эти акты входят в единую целенаправленную реакцию,
определяющую его поведение на сцене в данное время. Другие реакции, не
соответствующие доминантной установке, оказываются подавленными,
заторможенными. Любая форма трудовой деятельности представляет собою более или
менее сложную доминантную реакцию.
В зависимости от
текущих условий различные доминантные установки могут сменять друг друга
(переключение). Лишь в период смены доминантных установок и борьбы
«возникающей» и наличной «тормозимой» доминант может иметь место неполное,
абортивное проявление двух несовместимых, не включающихся в единую доминантную
систему реакций доминантного типа. При возникновении чесательного рефлекса у
бегущей собаки оба эти рефлекса — чесание и бег — становятся неполноценными, не
достигающими цели и, в конце концов, один из них — бег или чесание —будет
полностью подавлен, а другой нормально осуществится.
Принцип
доминанты в деятельности нервной системы включает в себя важнейшие
закономерности иррадиации возбуждений и межнейронных взаимодействий. Эти
закономерности суть следующие:
1. Наличие диффузной иррадиации импульсов
возбуждения в нервной системе. Эта закономерность обеспечивает
вероятностный характер рефлекторного реагирования, обусловленный богатством
коллатералей аксонов, благодаря которому каждый нейрон может влиять на тысячи
других нейронов в различных отделах нервной системы и входить с ними в те или
иные функциональные отношения. Возбуждение, возникающее в центральной нервной
системе, способно широко распространяться и достигать самых отдалённых её
участков не только при сильных раздражениях рецепторов или проводящих путей, но
и при обычных, слабых раздражающих воздействиях. Раздражение любого рецептора
или нерва может вызвать реакцию любого исполнительного органа.
2. Иррадиация импульсов возбуждения в сторону
нейронных групп, характеризующихся повышенной возбудимостью. Ещё в 1881 г.
Н.Е. Введенский показал, что тонически возбуждённый центр отвечает вспышкой
деятельности на раздражение любых чувствительных нервов. В 1896 г. он же,
описывая сопряжённые изменения возбудимости симметричных пунктов двигательной
зоны коры больших полушарий, отметил, что в течение известного времени после
раздражения определённого двигательного пункта коры раздражение других областей
коры вызывает эффект, характерный для только что раздражавшегося пункта.
А.А. Ухтомский в
серии блестящих экспериментов убедительно показал, что на иррадиирующие по
проводящим путям импульсы возбуждения первыми отвечают центры, характеризующиеся
повышенной возбудимостью (1911). Состояние повышенной возбудимости центров А.А.
Ухтомский связывал с развитием в них определённого уровня местного
стационарного возбуждения Н.Е. Введенского, но более низкого, чем уровень
стойкого местного возбуждения при развившемся парабиозе (запредельном
торможении). В этом смысле А.А. Ухтомский рассматривал состояние доминанты как
преддверие парабиоза.
Значительно
позже Ч. Шеррингтон и Дж. Экклс, характеризуя состояние центрального
возбуждения, связывали локальное возбуждение центров с повышением их
возбудимости и готовностью к импульсному разряду, возникающему с коротким
латентным периодом даже при слабых раздражениях (1931). Связь местного
подпорогового возбуждения нервных волокон с повышением возбудимости к этому
времени уже была подтверждена крупными представителями кембриджской
физиологической школы Эдрианом и Люкасом (1912). Уровень местного возбуждения
при повышении возбудимости характеризуется наличием менее глубокой
деполяризации нейронов по сравнению с уровнем местного возбуждения при снижении
и исчезновении возбудимости, связанными с глубокой деполяризацией. Местное
стационарное возбуждение нейронов, находящихся в состоянии доминанты,
характеризуется определенным уровнем их поляризации, связанным с повышением
возбудимости. Изучение доминантных состояний в моно- и полисинаптических
рефлекторных дугах показало, что в высоколабильных нейронах развитие
доминантных состояний связано со снижением их лабильности и поляризации
(классический механизм А.А. Ухтомского), а в малолабильных нейронах — с
повышением их лабильности и поляризации (Голиков, 1927, 1940).
Возникновение
доминантных состояний в нейронных группах, характеризующихся низким уровнем
лабильности, может быть связано не с деполяризацией и снижением лабильности, а,
напротив, с повышением уровня поляризации и лабильности. По-видимому,
формирование доминантных состояний при слабой анодической поляризации
поверхности коры, показанное В.С. Русиновым (1961, 1965) и подтвержденное Ф.
Мореллом (1961), связано с повышением лабильности и поляризации малолабильных
корковых элементов.
Исходя из
концепции местного возбуждения, выдвинутой Н.Е. Введенским в 1901 г., и
сопоставляя уровни местного возбуждения в нервных центрах при развитии
доминанты и возникновении пессимального торможения, А.А. Ухтомский пришёл к
выводу, что именно уровень местного возбуждения определяет возбудимость нервных
центров. По А.А. Ухтомскому, в силу высокой «впечатлительности» нервных клеток,
находящихся в состоянии местного возбуждения, и в зависимости от степени этой
впечатлительности одни и те же импульсы возбуждения могут вызывать либо
оптимальную реакцию возбуждения, связанную с возникновение импульсных разрядов,
либо пессимальную реакцию торможения.
Уровень местного
возбуждения, оценивавшийся Н.Е. Введенским и А.А. Ухтомским по величине
негативности возбужденной зоны, соответствует уровню деполяризации, причём
умеренная деполяризация (критический уровень деполяризации) и небольшой уровень
местного возбуждения характеризуются повышенной возбудимостью и возникновением
импульсных разрядов, а глубокая деполяризация и стойкое местное возбуждение —
исчезновением возбудимости, возникновением пессимального торможения Введенского
или запредельного торможения И.П. Павлова.
Следует
отметить, что термины «впечатлительность», «чувствительность», «реактивность»
клеток, волокон или их окончаний соответствуют лёгкости изменения
функционального состояния реагирующего субстрата, способности его к местной
реакции. Термин «возбудимость», предложенный Дюбуа Реймоном, обозначает
способность нервных и поперечнополосатых мышечных волокон отвечать на
раздражения бездекрементно распространяющимися импульсами возбуждения.
Реактивность (способность к местному ответу) и возбудимость (способность к
распространяющемуся ответу) изменяются неодинаково, неконгруентно.
Ещё в 1884 г.
Введенским было показано, что после исчезновения возбудимости тормозное
состояние участка нерва, подвергавшегося действию сильного постоянного тока,
может очень долго поддерживаться значительно более слабым током. Несмотря на
исчезновение возбудимости, чувствительность, реактивность нервных волокон
оказывается даже более высокой, чем в норме.
Аналогичные
факты были получены Л.Л. Васильевым, который показал, что сразу после
прекращения проведения альтерированный участок нерва обнаруживает высокую
чувствительность к слабейшим раздражениям. Можно ослабить или углубить
блокирование («пороговый парабиоз») слабыми раздражениями, например, пучком
ультрафиолетовых или инфракрасных лучей, или слабыми химическими воздействиями,
неэффективными для нормального нерва. Раздражимость, вначале повышающаяся
параллельно повышению возбудимости, в течение известного времени продолжает
повышаться и тогда, когда возбудимость уже снижается и исчезает. Современные
электрофизиологические исследования сделали очевидным тот факт, что местное
возбуждение может возникать и даже быть более выраженным при снижении и утрате
рефлекторной возбудимости. Медленная фоновая электрическая активность мозга и
локальные вызванные ответы на прямое и рефлекторное раздражения лучше выражены
при снижении и исчезновении рефлекторной возбудимости, при неглубоком наркозе,
сне, неглубоком запредельном торможении.
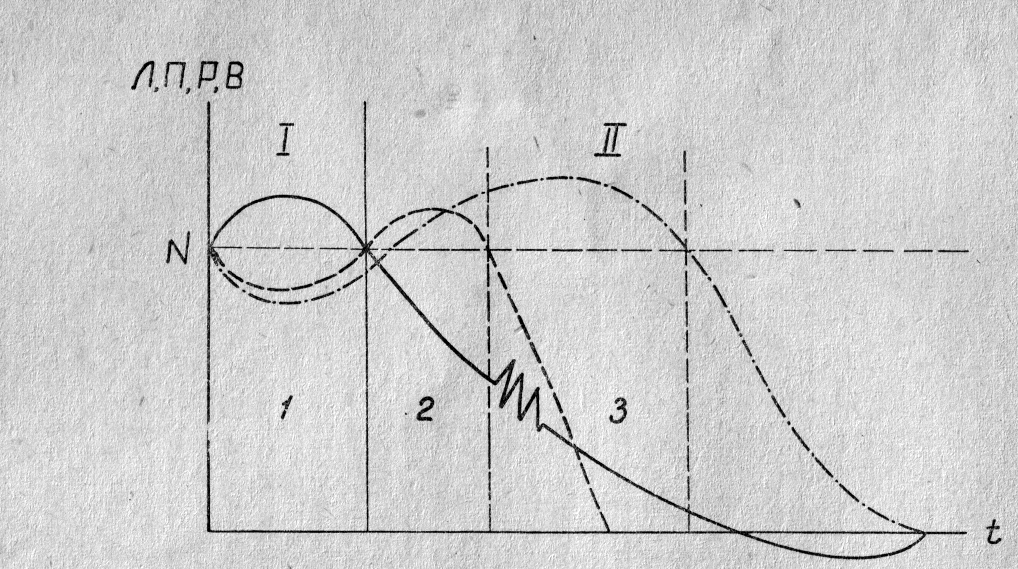
Рис.
5. Схематическое изображение изменений потенциала покоя возбудимости и
реактивности нервной ткани при длительном раздражении и развитии парабиоза.
По оси абсцисс — время действия
раздражителя, по оси ординат — изменения показателей функционального состояния
ткани. Сплошная линия — изменение поляризации и лабильности; прерывистая линия
— изменение возбудимости (способности к распространяющимся ответам);
прерывистая линия с точками — изменение реактивности (способности к местным
ответам). I — фаза повышения лабильности и
поляризации, II — фаза снижения лабильности и
поляризации. 1 — фаза первичного снижения возбудимости и реактивности, 2 — фаза
повышения раздражимости и возбудимости, 3 — фаза вторичного снижения
возбудимости и дальнейшего повышения, а затем вторичного снижения реактивности.
Наличие
медленной активности и облегчение возникновения локальных ответов при снижении
и исчезновении возбудимости требуют различать реактивность или раздражимость,
как более общее свойство клеток отвечать на раздражения изменением своего
состояния и местными реакциями и возбудимость как специализированную форму
раздражимости, как способность специализированных клеток отвечать на
раздражения распространяющимися нервными импульсами. Наибольшая реактивность и
наибольшая возбудимость клеток и волокон оказались связанными с определёнными,
но различными уровнями физиологической лабильности и поляризации этих клеток и
волокон (Голиков, 1933, 1950, 1952). При высоком уровне раздражимости легко
возникают местные возбуждения и местный автоматизм (местные пульсирующие
возбуждения и соответствующие им медленные потенциалы). При высоком уровне
возбудимости легко возникают бездекрементно распространяющиеся импульсы
возбуждения и импульсный автоматизм (множественные ритмические разряды нервных
клеток импульсами возбуждения) (рис. 5). В соответствии с закономерностями
эволюционной физиологии, показанными Л.А. Орбели (1949), филогенетически более
новая форма реагирования импульсными разрядами развивается на базе более
древней формы реагирования местными реакциями.
Привычные
представления о возбуждении как появлении или усилении импульсации и о
торможении как о подавлении импульсных разрядов оказались справедливыми лишь
для системных реакций нервных центров. Клеточные реакции, в частности местная
электрическая активность и местные реакции на раздражения, не только
сохраняются в течение некоторого времени после прекращения проведения импульсов
возбуждения, но даже усиливаются в первые моменты после наступления
блокирования проведения. Длительная сохранность местных ответов после
прекращения проведения при развитии парабиоза в нашей лаборатории показана М.И.
Сологубом (1957), а в лаборатории Д.Н. Насонова — Г.Н. Можаевой (1958) в
условиях регистрации локальных ответов в месте раздражения. Локальные
возбуждения возникают в широком диапазоне изменений уровня функционального
состояния клеток и их отростков, и если применять термины «возбуждение» и
«торможение» при анализе клеточных реакций, то нужно связывать возбуждение с
повышением раздражимости и обмена веществ клеток, а торможение — со снижением
их раздражимости и обмена веществ. В зависимости от исходного уровня функционального
состояния нервных клеток и волокон раздражающие воздействия, в частности сами
нервные импульсы, могут либо снижать, либо повышать раздражимость и
возбудимость. При высоком уровне поляризации и лабильности изменения
возбудимости (способности к бездекрементно проводящимся ответам) и изменения
реактивности (способности к местному ответу) протекают параллельно, при
значительно сниженном уровне лабильности и поляризации изменения возбудимости и
реактивности протекают в диаметрально противоположном направлении (рис. 6).
Характер ответной реакции нейронов на иррадиирующие в нервной системе импульсы
возбуждения зависит в значительной степени от уровня реактивности нейронов.
Повышение реактивности нейронов приводит к тому, что синаптические воздействия,
вызывавшие ранее лишь трофический эффект повышения поляризации и лабильности и
снижение возбудимости, теперь будут действовать в сторону деполяризации,
повышения возбудимости и возникновения импульсных разрядов нейронов. Ранее
оптимальное, возбуждающее действие более значительной импульсации при
значительном повышении реактивности нейронов может вызвать развитие глубокой
деполяризации, снижение возбудимости и возникновение пессимального,
запредельного торможения. Фоновая афферентная импульсация, оказывающая на
высоколабильные нейроны лишь положительно-трофическое действие, связанное с
повышением их лабильности и поляризации и снижением возбудимости, при
значительном повышении реактивности нейронов будет вызывать возбуждающее
действие, связанное с умеренной деполяризацией, некоторым снижением лабильности
и повышением возбудимости. Эта импульсация начнёт подкреплять возбуждение
доминирующей группы нейронов.
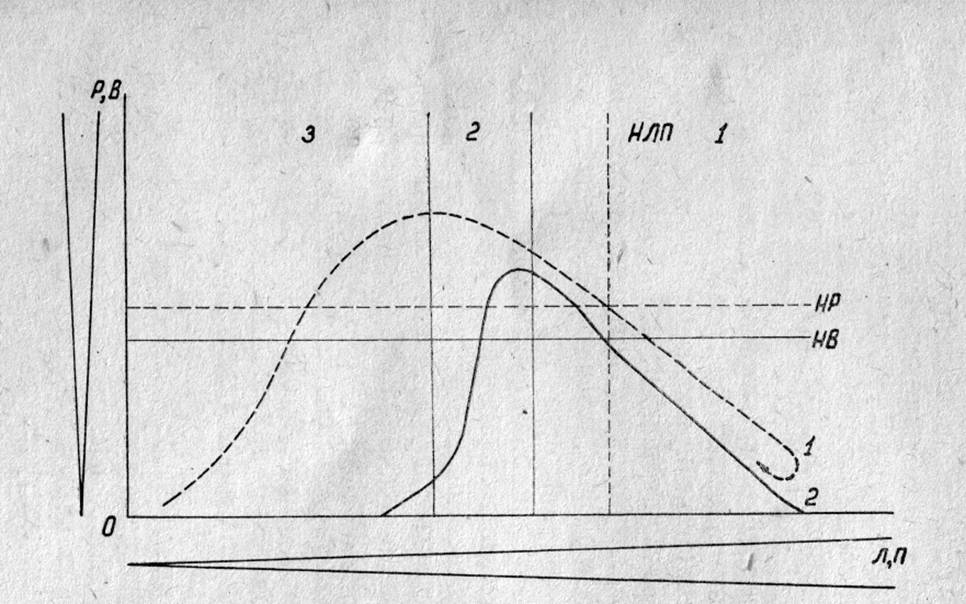
Рис.
6. Схематическое изображение зависимости раздражимоти и возбудимости нервных
клеток и волокон от уровня их поляризации и лабильности.
По оси абсцисс — возрастающее
значение лабильности и поляризации (Л, П), по оси ординат — возрастаюшее
значение раздражимости (Р) и возбудимости (В). НР — нормальный уровень
раздражимости, НВ — нормальный уровень возбудимости, НЛП — нормальный уровень
лабильности и поляризации. Сплошная линия — изменение возбудимости, прерывистая
линия — изменение раздражимости (реактивности). 1 — зона одновременного
повышения реактивности и возбудимости при умеренном снижении лабильности и
небольшой деполяризации, 2 — зона повышения раздражимости и снижения
возбудимости при дальнейшем снижении лабильности и уровня поляризации, 3 — зона
отсутствия возбудимости и снижения реактивности при ещё более значительном
снижении лабильности и поляризации.
3. Явления сопряжённого торможения.
Центры, находящиеся в состоянии повышенной возбудимости, не только первыми
отвечают на иррадиирующие в нервной системе импульсные возбуждения своим
рабочим эффектом, но и оказывают сопряжённо тормозящее действие на другие
центры. По мере формирования системы совозбуждённых центров сопряжённое
торможение других центров всё возрастает. По наблюдениям А.А. Ухтомского,
иррадиирующие в нервной системе импульсы возбуждения подкрепляют состояние
возбуждения в центрах с повышенной возбудимостью и углубляют торможение в
центрах с более низкой и со сниженной возбудимостью. Для влияния очагов
повышенной возбудимости на другие центры оказались характерными феномены
сопряженного изменения возбудимости, описанные для двигательной зоны коры
больших полушарий Н.Е. Введенским в 1897 г., и аналогичные им явления
реципрокного торможения, изученные Ч. Шеррингтоном в рефлекторных дугах
спинного мозга (1906). Явления отрицательной индукции по одновременности
оказались широко представленными не только в сложившихся рефлекторных дугах, но
и в различных «временных» рабочих, констелляциях нервных центров.
Механизм
различного влияния иррадиирующих в нервной системе импульсов возбуждения на
различные группы нейронов до настоящего времени остаётся дискуссионным. Ряд
исследователей связывает тормозящие и возбуждающие влияния иррадиирующих в
нервной системе нервных импульсов лишь со специализацией межцентральных связей
и синаптических аппаратов. Однако общие закономерности раздражений,
динамичность межцентральных взаимоотношений и возможность перестройки обычных
иннервационных взаимоотношений даже в сложившихся рефлекторных дугах
показывают, что в этих влияниях важную роль играет смена специфического для
живых образований начального положительно-трофического, первично-тормозного
эффекта действия раздражений последующим возбуждающим, а затем вторично
тормозящим их действием в результате закономерного изменения уровня
функционального состояния реагирующих нейронов. Различная реактивность разных
групп специализированных нейронов обусловливает возникновение различных реакций
на одну и ту же импульсацию. Трёхфазность рефлекторной реакции, часто
наблюдающуюся при сильных и длительных раздражениях рефлекторных дуг, иногда
объясняют тем, что более высокая возбудимость специализированных тормозных
систем обусловливает начальные тормозные эффекты, последующее возбуждение
связывается с последующей активацией специфических возбуждающих структур, а
вторичное торможение — с развитием пессимума Введенского под влиянием
чрезмерного действия тех же нормально возбуждающих структур. В нейронных реакциях
на афферентную и межцентральную импульсацию имеет место как суммарный результат
влияния специализированных синаптических аппаратов, так и закономерности
неспецифической реакции фазного изменения функционального состояния нейронов,
возникающей при любых раздражающих воздействиях. Эффект сопряженного
торможения, характерный для обычного действия диффузной асинхронной
импульсации, может подчёркиваться специфическим гиперполяризующим действием
специализированных тормозных структур в тех случаях, когда эти структуры
оказываются более реактивными, чем специализированные возбуждающие структуры.
Материалы эволюционной физиологии, показывающие развитие специфических эффектов
действия медиаторов из первичного неспецифического их действия, позволяют
рассматривать специфические эффекты специализированных синаптических структур
как подчёркивание и затягивание, преимущественное выражение одной из фаз
неспецифической реакции на раздражение. Эффекты активации специализированных
синапсов зависят от уровня функционального состояния реагирующих клеток. В
зависимости от уровня поляризации, лабильности и реактивности нейрона
трофическое действие медиатора может смениться тормозящим его действием,
тормозящее действие медиатора может смениться его возбуждающим действием. Для
возникновения сопряжённого торможения нейронных групп, не вошедших в состав
доминирующей констелляции нервных центров, необходимо наличие определённого
уровня поляризации и реактивности нейронов и, возможно, определенная степень
активации их специализированных синаптических аппаратов. По данным А.А.
Ухтомского, сопряжённое торможение нервных центров, не вошедших в состав
доминирующей констелляции, оказывается более интенсивным и более выраженным в
более лабильных рефлекторных дугах и менее выраженным — в менее лабильных
рефлекторных дугах, даже при более близком территориальном их расположении по
отношению к доминантному очагу. При ухудшении функционального состояния нервной
системы, при упадке лабильности и поляризации нейронов сопряжённое торможение
ослабевает и даже сменяется повышением возбудимости и возникновением
возбуждения в ранее сопряжённо тормозившихся центрах. Явления сопряжённого
торможения при формировании сложных доминантных установок могут быть связаны
также с формированием избирательного отношения нейронов к импульсации
различного ритма (различного кода). При импульсации, характеризующейся
определенным ритмом, имеет место возникновение суммации супернормальности и
повышение возбудимости нейронов именно для этой импульсации. При импульсации, характеризующейся
другим ритмом, напротив, имеют место суммация субнормальности и снижение
возбудимости. Импульсация данного ритма вызывает торможение нейрона.
4. Явления синхронизации активности нейронов,
входящих в состав доминирующей констелляции. Ещё в 1924 г. А.А. Ухтомский
выдвинул положение, по которому каждая доминирующая система нервных центров
характеризуется единым ритмом активности. Позже, развивая свои представления о
механизмах доминантных реакций при анализе речевой деятельности и используя накопившиеся
данные о явлениях усвоения нервными центрами ритма раздражения, он стал
связывать феномены вовлечения (рекрутирования) и феномены выключения
(демобилизации) тех или иных нейронных групп (центров) при формировании
доминирующих констелляций со способностью этих нейронных групп усваивать единый
ритм активности ведущего центра. В отличие от первоначальных представлений об
усвоении ритма, связывавших его только с перестройкой активности нейронов на
более высокий ритм деятельности при повышении их лабильности, А.А. Ухтомский
стал рассматривать процесс сонастраивания центров на единый ритм активности как
результат снижения лабильности более лабильных нейронных групп и повышения
лабильности низколабильных нейронных групп, входящих в состав доминирующей констелляции.
Это представление об усвоении ритма подкреплялось данными, по котором усвоение
ритма связано с повышением возбудимости и характерно для доминантных состояний
(Голиков, 1927, 1930). Синхронизация активности нейронных групп, входящих в
состав доминирующих констелляций нервных центров, формируется постепенно. В
начале развития доминантных состояний и при элементарных доминантах имеет место
лишь синхронизация темпа активности. Тонкое сонастраивание ритмической
активности, усвоение не только темпа, но и ритма возникают значительно позже,
при повторном возникновении деятельных состояний. Одновременно постепенно
развивается и способность к репродукции усвоенного ритма. В центрах формируются
достаточно прочные специфические следы предыдущих раздражений.
Усвоение и
репродукция ритма активности, по-видимому, связаны с формированием
специфических следов раздражений, обеспечивающих возникновение избирательно
высокой возбудимости для импульсации определённого ритма.
Роль процессов
усвоения ритма и следовых процессов при образовании доминантных установок
становится все более ясной по мере накопления данных об изменениях ритмической
активности нейронов и нейронных групп при различных функциональных состояниях
мозга. Явления синхронизации электрической активности нейронных групп в
различных корковых полях и подкорковых образованиях при формировании системных
реакций нервных центров, впервые показанные в лабораториях М.Н. Ливанова и В.С.
Русинова, сейчас неоднократно подтверждены.
В феноменах
усвоения ритма раздражений нервными центрами различается усвоение ритма в
реакциях типа местных ответов и в реакциях типа импульсных разрядов. Феномены
усвоения ритма, выражающиеся в синхронизации местных ответов и местной
активности, по нашим данным, сочетаются с повышением реактивности нейронов и
наиболее выражены при определённом невысоком уровне их лабильности и
поляризации. Феномены усвоения ритма, выражающиеся в синхронизации импульсных
разрядов и импульсной активности, наиболее выражены при более высоком уровне
поляризации и лабильности нейронов. Нами (1949), а позже Фессаром (1961)
синхронизация активности соседних нейронов друг с другом была названа
внутренней синхронизацией, а синхронизация активности нейронов с ритмом
приходящей импульсации — внешней синхронизацией. Феномены восстановления
активности в прежнем, навязанном предыдущей стимуляцией ритме нами обозначены
термином «репродукция усвоенного ритма» и связываются с длительным сохранением
специфических следов раздражений нейронами высших отделов нервной системы в виде
избирательного повышения возбудимости этих нейронов к импульсации определенного
ритма. При формировании доминантной констелляции нервных центров по А.А.
Ухтомскому вначале имеет место генерализованное возбуждение ряда центров, в
доминантную констелляцию вовлекаются и не необходимые центральные клеточные
группы. По мере повторного осуществления доминирующего акта ненужные для этого
акта нейронные группы затормаживаются, выключаются из работы, реализация
доминантного акта становится экономной. Так же, как при образовании условного
рефлекса, при формировании доминанты начальный этап избыточного вовлечения
нейронных групп (генерализованная реакция) по мере упрочения сменяется этапом
концентрирования активности нейронов только в тех клеточных группах, которые
необходимы и достаточны для реализации данного доминирующего акта. Вместе с тем
диффузная реактивность доминирующей констелляции центров к различным
раздражениям в процессе повторения рабочего акта постепенно сменяется
избирательным ответом этой констелляции только на раздражения, непосредственно
формирующие данную доминанту. В элементарных доминантных реакциях единство
действия констелляции совозбуждённых центров осуществляется в порядке усвоения
единого темпа активности. Тонкое сонастраивание активности нейронов на единый
ритм связано с формированием специфических следов раздражений и возникновением
избирательно высокой отзывчивости нейронов к стимуляции определённого ритма.
Если нейронные группы, способные усвоить и сохранить данный вид активности, вовлекаются
в функциональную констелляцию, то другие группы, не способные усвоить этот
ритм, затормаживаются и выключаются из доминирующей констелляции. Некоторые
формы центрального торможения и, по-видимому, внутреннее (условное) торможение
И.П. Павлова связаны с явлениями воспроизведения усвоенного ритма и с
формированием длительно сохраняющихся специфических следов раздражения.
Н.Е. Введенский,
различал положительное, и отрицательное взаимное усиление (корроборацию)
действия слабых, в отдельности неэффективных раздражений и способность слабого,
ранее неэффективного раздражения длительно поддерживать реакцию возбуждения или
торможения, возникшую при действии сильных раздражений. Рефлекторное
возбуждение, вызванное совместным действием двух неэффективных в отдельности
раздражителей, может длительно поддерживаться одним из этих раздражений.
Торможение, созданное сильным раздражением, может поддерживаться раздражителем,
значительно более слабым (методика минимальной поляризации). Явления
внутреннего торможения, характерные для формирования условного рефлекса и
сложных доминирующих системных реакций нервных центров, отличаются от других
форм торможения постепенностью развития и связаны не только с гиперполяризацией
нейронов, но и с избирательным «отрицательным» реагированием на импульсацию
определённой модальности. В этом существенное отличие постепенно формирующегося
внутреннего (условного) торможения от безусловного внешнего торможения, обычно
также связанного с явлениями гиперполяризации. Давние электрофизиологические
наблюдения Верцара (1928) и Гассера (1935) показали, что ритмическая стимуляция
в зависимости от её частоты и длительности может создавать в нервных волокнах
явления гиперполяризации или явления деполяризации в порядке суммации следовых
изменений (субнормальности или супернормальности), остающихся после каждого
импульса. К этим феноменам близки явления посттетанического усиления
(потенциирования), связанные с гиперполяризацией концевых аксонов, и явления
пресинаптического торможения, напротив связанного с глубокой деполяризацией
этих разветвлений.
5. Экзальтационные и тормозные последействия в
рефлекторных реакциях и длительная сохранность специфических следов раздражений
в нейронах высших отделов нервной системы. Эволюция нервной деятельности и
формирование новых рефлекторных реакций обусловлены длительным сохранением
следов раздражений нервными клетками. Еще И.М. Сеченов различал два типа следов
— кратковременные, возникающие по прекращении раздражения и скоро затухающие, и
следы, длительно сохраняющиеся, обусловливающие явления памяти. Простые следы
представляют собою последействия — однозначные и контрастные с реакцией на
раздражение — и соответствуют явлениям последовательной центральной индукции.
После прекращения раздражения вызванная им рефлекторная реакция может
продолжаться в течение некоторого времени (положительный след) и лишь затем
временно сменяться противоположным эффектом. Такая реакция может быть названа
задержанным последовательным контрастом. Контрастное изменение знака реакции на
раздражение может наступить сразу после прекращения этого раздражения. Такая
следовая реакция может быть названа непосредственным последовательным
контрастом. Примером такого непосредственного контраста может служить «рефлекс
Сеченова». Чрезмерно сильное раздражение вызывает тормозное состояние, которое
сменяется экзальтационной реакцией сразу после прекращения тормозящего
раздражения. Простые следовые реакции или последействия того или иного знака
затухают волнообразно, этапы повышения и снижения возбудимости нейронов
становятся всё менее выраженными и через несколько минут после прекращения
раздражения простые следы полностью исчезают.
В любой
рефлекторной дуге, изменяя лишь величину раздражения, можно вызвать как
состояние возбуждения, так и состояние торможения и простые следовые реакции
типа непосредственного или задержанного контраста (Голиков, 1927).
Простые следовые
реакции подразделены на четыре типа, которые сменяют друг друга в зависимости
от величины раздражения и изменения состояния реагирующих нейронов. Первый тип
— экзальтационное последействие (положительный след, задержанная
последовательная отрицательная индукция). После прекращения раздражения
рефлекторное возбуждение нейронов продолжается в течение некоторого времени и
лишь затем подавляется. Второй тип — торможение вслед за возбуждением
(отрицательный след, непосредственная последовательная отрицательная индукция).
Тотчас по прекращении раздражения состояние возбуждения сменяется временным
тормозным состоянием. Третий тип — экзальтация вслед за торможением
(непосредственная последовательная положительная индукция), тотчас после
прекращения тормозящего раздражения торможение сменяется вспышкой возбуждения.
Четвёртый тип — тормозное последействие (задержанная положительная
последовательная индукция). По прекращении тормозящего раздражения тормозное
состояние длится в течение некоторого времени и лишь затем сменяется
возбуждением. Для простых следовых реакций (последействий) характерно
неизбирательное, имеющее место для любых раздражителей следовое изменение
рефлекторной возбудимости — снижение и исчезновение её при следовых торможениях
и повышение её при следовых экзальтациях. Специализация нейронов и синапсов
обусловливает тот факт, что в качестве первичной реакции легко возникает и
прочно удерживается либо реакция рефлекторного возбуждения и соответствующие ей
следовые реакции зкзальтационного последействия и торможения вслед за
возбуждением, либо реакция рефлекторного торможения и соответствующие ей
следовые реакции тормозного последействия и экзальтации вслед за торможением.
Два других типа следовых реакций возникают лишь при значительном увеличении
раздражений или при значительном изменении функционального состояния
реагирующих нейронов. Так, при раздражении «возбуждающих» афферентных волокон
легко возникают и легко переходят друг в друга следовые реакции типа
экзальтационного последействия и торможения вслед за возбуждением. При
раздражении «тормозящих» афферентных волокон (то есть волокон, связанных со
специфическими тормозящими синапсами) легко возникают и переходят друг в друга
реакции тормозного последействия и экзальтации вслед за торможением.
Специализация нейронов и синапсов значительно расширяет диапазон специфически
возбуждающего или специфически тормозящего действия нервных импульсов.
Специфические
следы, лежащие в основе памяти, связаны с избирательным повышением возбудимости
нейронов к импульсации определённого ритма, к «кодированной» импульсации.
Возможно, что избирательное повышение реактивности и возбудимости для
определённого ритма раздражений («усвоение ритма раздражений и способность к
репродукции усвоенного ритма») связано с перестройкой молекул нативного белка и
матричных молекул нуклеиновых кислот в реагирующих нейронах. Эта перестройка и
увеличение пластического обмена могут привести к усилению развития
специализированных возбуждающих и тормозящих связей между различными
специализированными нейронными группами и к облегчению определённых
взаимоотношений между этими группами. Наличие длительно сохраняющихся
специфических следов и появление избирательного реагирования на импульсацию
определённой модальности обусловливают возможность сигнального действия
раздражителей и процессы анализа (сопоставление текущих раздражений со следами
предыдущих, «узнавание» раздражителей).
Наличие длительно
сохраняющихся следов раздражения обеспечивает формирование новых функциональных
констелляций активных нейронных групп, осуществляющих новые рефлекторные акты и
определяющих поведение человека и животных.
Длительная
сохранность специфических следов раздражений характерна для нейронов высших
отделов нервной системы, у высших животных — для нейронов коры. Высшие отделы
нервной системы, кора больших полушарий высших животных и человека представляют
собою специализированный аппарат, осуществляющий хранение следов предыдущих
раздражений, выработку новых рефлекторных реакций на раздражающие воздействия
среды и предупредительную ориентировку в среде. Животное или человек, опираясь
на имеющийся опыт (следы прежних воздействий) и используя приобретённые навыки
и умения (выработанные новые рефлекторные акты) по поводу сигнальных
раздражений, поступающих из дистантных рецепторов (зрительного, слухового,
обонятельного), может совершить ряд предупредительных действий, необходимых для
того, чтобы избежать опасности при добывании пищи и т. д. Как при недоразвитии
коры больших полушарий, так и при органических или функциональных её поражениях
на первый план выступают нарушения памяти, способности к обучению и к
целенаправленной деятельности.
Способность к
длительному сохранению специфических следов раздражений нейронными группами
высших отделов нервной системы лежит в основе сигнального действия
раздражителей, связанных с процессами анализа и образования условных рефлексов.
Всё развитие и совершенствование нервной системы в первую очередь обусловлено
развитием способности нейронов её высших отделов длительно сохранять
специфические следы всё более сложных раздражающих воздействий.
6. Системность рефлекторных актов. Каждый
рефлекс осуществляется комплексом нейронных групп, расположенных в различных
отделах мозга, но объединённые в одну систему, констелляцию, деятельность
которой поддерживается циклами межцентральных импульсов и циклами импульсов
между центрами и периферическими исполнительными аппаратами. Группа одновременно
возбуждающихся нейронов образует систему, констелляцию, осуществляющую тот или
иной рефлекторный акт, синхронизирует свою деятельность с ритмом и темпом
активности ведущих нейронных групп, расположенных в высших отделах мозга и
способных длительно сохранять следы раздражений. При вхождении тех или иных
групп нейронов в другие функциональные констелляции их активность
перестраивается на другой ритм, характерный для данной, другой констелляции,
возглавляемой другими ведущими нейронными группами высших отделов мозга. По
мысли А.А. Ухтомского, сложившиеся рефлекторные дуги низших отделов нервной
системы могут использоваться в разных комбинациях и по разным поводам её
высшими отделами. Прежние относительно простые рефлекторные акты,
осуществляемые сложившимися специализированными рефлекторными дугами, частично
тормозятся, частично перекрываются новыми рабочими применениями. «Общий
исполнительный путь приобретает новые и новые адекватные поводы и рецептивные
аппараты, а также качественно новое значение в составе, новых рефлекторных
актов. Всё новые и новые адекватные поводы, совершенно не существовавшие для
низших этажей в их отдельности, по мере развития нервной системы получают
доступ и стимулирующее значение для прежнего исполнительного пути с его
механизмами» (А.А. Ухтомский, 1942). Развиваются и новые спрямлённые пути
(пирамидная система млекопитающих), непосредственно связывающие высшие отделы
нервной системы с эфферентными исполнительными нейронами. «Когда в нижележащих
этажах успели выработаться определённые рефлексы и рефлекторные дуги с точно
приуроченными к ним адекватными рецепторами, следующий более высокий этаж
отправляется в своих рефлекторных работах от этих предыдущих выработок, как от
данных заранее, используя их в составе своих более высоких рефлексов уже по
новым адекватным поводам, часто независимым и даже вопреки стимулам с прежних
рецепторов низших этажей. Если эти последние стимулы из прежних рецепторов были
до сих пор обязательными, т. е. безусловными для рефлексов низших этажей, то с
включением в состав рефлексов высших этажей и качественно нового содержания
прежний аппарат и эффект приобретают существенно новое значение, содержание и
функциональное качество, становясь независимыми от прежних адекватных, т. е.
безусловных поводов вследствие того именно, что вступают стимулы с более
высоких этажей» (А. А. Ухтомский, 1942).
Давно были
известны факты, показывавшие, что при наличии одновременного возбуждения двух
нервных центров возбуждение одного из них может поддерживаться нервными
импульсами, иррадиирующими из другого, совозбуждённого центра (явления
корроборации Введенского, 1881; проторения пути Экснера, 1882; схема взаимного
связывания двух совозбуждённых центров Экклса и Гранита, 1929).
При выработке
условного рефлекса одним из необходимых условий образования временной связи
между группами промежуточных нейронов в зонах коркового представительства
условного и безусловного раздражителей является хотя бы частичное совпадение во
времени состояний возбуждения в этих центрах. Тогда в соответствии с законом
иррадиации возбуждений эти нейронные группы, характеризующиеся повышенной
возбудимостью, будут взаимно подкреплять свое возбуждение, окажутся взаимно
связанными. Однако для репродукции системной реакции одновременного возбуждения
центров условного и безусловного раздражителей по поводу действия одного
условного раздражителя необходимо длительное сохранение состояния повышенной
возбудимости в этих центрах. Простые следы раздражений затухают очень быстро,
повышение возбудимости, лежащее в основе условнорефлекторных реакций, весьма
длительно и имеет избирательный характер. Поэтому не простые следы, не
последействия лежат в основе выработки условного рефлекса.
Другим важнейшим
условием формирования сложных системных реакций вроде условного рефлекса или динамического
стереотипа является длительное сохранение специфических следов раздражений
нейронными группами. Эти следы закрепляются и упрочиваются при повторном
возникновении системной реакции в результате одновременного действия условного
и безусловного раздражителей (обучение). Третьим условием, необходимым для
формирования системной реакции нейронов, является наличие определённого
оптимального уровня функционального состояния мозга. Обеспечивается этот
уровень определённой интенсивностью обмена веществ, оптимальным притоком
раздражений из рецепторов, импульсацией из высших симпатических центров (Л.А.
Орбели, 1939) и из ретикулярной формации мозгового ствола и межуточного мозга
(Моруцци и Мегоун, 1949).
В своём развитии
условный рефлекс проходит начальную фазу генерализации, во время которой
условнорефлекторная реакция может быть вызвана не только данным условным, но и
близкими к нему раздражителями, и последующую фазу концентрации или
закрепления, когда рефлекс вызывается только данным, адекватным для него
условным раздражителем. Электрофизиологические исследования механизма
образования условных рефлексов подтвердили данные, полученные с помощью
классической методики, и позволили обнаружить ряд новых фактов. Уже при первых
электрофизиологических исследованиях формирования условных рефлексов (М.Н.
Ливанов и др., 1934, 1951) было показано наличие генерализованных
(общекорковых) и очаговых (локальных) изменений биоэлектрической активности
коры больших полушарий в процессе выработки этих рефлексов. При укреплении
условного рефлекса генерализованные, общекорковые изменения электрической
активности исчезали. Выяснилось, что корковые, диффузные изменения
электрической активности обусловлены главным образом неспецифическими
активирующими влияниями из ретикулярной формации мозгового ствола и межуточного
мозга, соответствующими адаптационно-трофическим влияниям, описанным Л.А.
Орбели с сотрудниками. Постоянство усиления активности ретикулярной формации в
начале выработки условного рефлекса и наличие условнорефлекторных изменений
электрической активности коры побудило некоторых авторов рассматривать
ретикулярную формацию как образование, в котором происходит замыкание временных
связей (Фессар и Гасто, 1956; Гасто, 1957). Однако накопилось много данных,
которые показывают, что при образовании условного рефлекса временно и в
известной последовательности вовлекаются в активность не только образования
ретикулярной формации, но и ряд других центральных образований, имеющих
типичную ядерную или корковую структуру, например вентральные ядра таламуса и
извилина морского коня. При выработке условного рефлекса десинхронизация
фоновой электрической активности коры, возникавшая при действии условного
раздражителя, вскоре сменяется реакцией появления синхронизированных медленных
потенциалов в ответ на действие того же раздражителя. В это время появляется
генерализованная условнорефлекторная реакция. Наконец, действие условного
раздражителя вызывает лишь реакцию очаговой десинхронизации и
дифференцированный чёткий условный рефлекс.
Л.Г. Трофимов с
сотрудниками обнаружили, что при упрочении условного рефлекса ретикулярная
формация перестает отвечать на условное раздражение генерализованным
активирующим действием. Аналогичные данные были получены Мореллом при изучении
клеточных разрядов, возникающих в различных отделах мозга при выработке
условного рефлекса. В начале выработки условного рефлекса клеточные разряды
возникали в проекционных зонах условного и безусловного раздражителей, в
гиппокампе и в ретикулярной формации. Затем при укреплении рефлекса клеточные
разряды возникали в проекционных зонах и переднем вентральном ядре таламуса, но
отсутствовали в ретикулярной формации ствола и в дорзальном гиппокампе. Эти
данные не позволяют усматривать в ретикулярной формации специальный механизм замыкания
временной связи. Несомненно лишь то, что наличие тонизирующих, активирующих
влияний из ретикулярной формации необходимо как для выработки, так и для
нормального осуществления условных рефлексов. Роль адаптационно-трофических
симпатических влияний в условнорефлекторной деятельности ранее отмечалась Л.А.
Орбели (1932, 1939). Нет оснований также связывать активирующее влияние только
с ретикулярной формацией. Активирующее влияние осуществляется не только
неспецифическими (ретикулярными), но и специфическими таламо-кортикальными
(афферентными) волокнами (Л.Г. Трофимов и др., 1958; Чанг, 1958).
Внутриклеточное
отведение потенциалов от различных зон коры головного мозга показало наличие
импульсного автоматизма нейронов при возбуждении и прекращение или подавление
их импульсных разрядов при торможении. Изменения поверхностной
электрокортикограммы не всегда коррелировали с изменением разряда нервных
клеток. Поверхностная десинхронизация обычно связана с усилением клеточных
разрядов, однако в некоторых клетках наблюдались подавление и прекращение
разрядов. Данные Морелла и Ливанова показывают наличие корреляции между
разрядами клеток и поверхностной десинхронизацией и корреляции между
подавлением разрядов клеток и поверхностной синхронизацией медленных потенциалов
при сочетании глубинного и поверхностного отведения потенциалов. Такие же
данные были получены в нашей лаборатории Т.Ш. Хапажевым (1963) и Р.С. Мнухиной
(1964).
В комплексе
условий, необходимых для образования временной связи между центрами, Фессар
выделяет необходимость конвергенции нервных импульсов, их взаимодействия и
длительного сохранения вызванных ими изменений. Конвергенция и взаимодействие
нервных импульсов в общих путях, а также определённый оптимальный уровень
функционального состояния мозга необходимы и для возникновения безусловных
системных реакций мозга. Для условных рефлексов и для репродукции доминантных
реакций, кроме того, необходима длительная сохранность специфических следов
раздражений, обозначаемая как «скрытые следы» (И.М. Сеченов), «высокая
запечатлеваемость нейронов» (И.П. Павлов), «сохранение следов прошлого»
(Фессар), «удерживание», «пластичность». Именно длительная сохранность
специфических следов раздражения и обусловливает тот факт, что один условный
раздражитель способен вызвать всю системную реакцию, ранее вызывавшуюся
комплексом «условных» и «безусловных» раздражений. Джаспер и Морелл
рассматривают образование временной связи и сигнальное значение условного
раздражения как возникновение специального механизма, облегчающего передачу
кодированных во времени и пространстве серий импульсов возбуждения по
специфическим и неспецифическим путям. Это представление созвучно с
представлением А.А. Ухтомского о специфическом ритме активности каждой
доминирующей констелляции нервных центров и о длительной сохранности следов
раздражения в этих центрах. На основании данных сравнительной физиологии и
сведений о возможности выработки примитивных условных рефлексов у животных,
лишённых коры больших полушарий, можно считать установленным, что замыкание
временных связей может происходить на тех уровнях центральной нервной системы,
клетки которых способны достаточно долго сохранять специфические следы
раздражений. Примитивные временные связи могут замыкаться и в ретикулярной
формации. Вместе с тем ретикулярная формация регулируется корой головного мозга
и используется как эффекторный аппарат корковых иннервации. П.К. Анохин (1958)
получил убедительные факты, показывающие подчинённый характер участия
ретикулярной формации в ряде сложных рефлекторных актов, связанных с тонкими
дифференцировками. В системных реакциях ретикулярная формация, как и другие
отделы нервной системы, может играть как роль инициатора, так и роль
исполнителя в зависимости от структуры и фазы развития данной реакции. Однако
тонкая и сложная динамика нервных процессов при высшей нервной деятельности
обусловлена возникновением и укреплением связей между кортикальными участками
тех или иных констелляций нервных центров. Сведения о характере, путях и
механизме иррадиации нервных импульсов, полученные с помощью
электрофизиологической методики в сочетании с перерезкой определенных путей,
показали наличие двух способов взаимной связи корковых полей: ассоциативными
волокнами в самой коре и корково-подкорково-корковыми путями (Пурпура и Грундфест,
1956, и др.). Нужно считать недоразумением представление о том, что взаимосвязь
корковых полей осуществляется только через подкорковые образования.
*
* *
Выдвинутая А.А.
Ухтомским концепция функциональной констелляции нервных центров, осуществляющих
ту или иную системную реакцию, в настоящее время получает всё новые
подтверждения и всё более используется, часто без упоминания её автора.
Накапливающиеся данные подтверждают системный характер сочетанной деятельности
нервных элементов различных этажей нервной системы при осуществлении как
условных, так и безусловных рефлексов и наличие общих закономерностей этой
деятельности. При электрофизиологическом изучении механизма возникновения
системных реакций мозга отмечены закономерные изменения степени синхронизации
медленной и быстрой ритмической активности нервных клеток и подтверждены старые
наблюдения Бергера, по которым изменения медленных потенциалов, соответствующих
местным возбуждениям, менее коррелируют с динамикой высшей нервной
деятельности, чем изменения быстрых потенциалов, связанных с возникновением
импульсных разрядов. Внутриклеточное отведение потенциалов из различных отделов
мозга подтверждает связь динамики формирования системных реакций с изменениями
в характере импульсных разрядов и следовыми процессами. Сделано в этом
направлении ещё мало, но уже полученные факты позволяют подчеркнуть важность
представлений А.А. Ухтомского о роли процессов усвоения ритма (сонастраивания)
и об экономизирующем значении синхронизации при формировании условных и
безусловных системных реакций.
Общие
закономерности воздействия раздражений определяют характер реакции нейронов на
синаптические и непосредственные раздражения и обусловливают возникновение
состояния возбуждения или торможения. Специализация рецепторов, нейронов и
синапсов обеспечивает относительное постоянство и надежность рефлексов.
Развитие специализированных структур в различных отделах мозга экономизирует и
оптимизирует рефлекторную деятельность. Чувствительные, ассоциативные и
исполнительные (двигательные и вегетативные) группы нейронов в различных
комбинациях образуют рабочие системы, констелляции, осуществляющие различные
сложные акты. Усвоение ритма раздражений (синхронизация активности нейронов
друг с другом и с ритмом приходящих раздражений) — один из важных механизмов
рекрутирования и выключения нейронных групп при формировании функциональных
констелляции нервных центров, осуществляющих рефлекторные акты. Репродукция
системных реакций и возникновение новых, рефлекторных актов тесно связаны со
способностью нейронных групп длительно сохранять следы предыдущих раздражений.
Эти следы, определяющие явления памяти, способность к обучению, способность к
предупредительной ориентировке в среде и развитие, и совершенствование нервной
деятельности, характеризуются избирательным повышением реактивности нейронных
групп к кодированной импульсации. Вскрываемые при физиологических исследованиях
нервной деятельности механизмы работы мозга используются в технике при
конструировании сложных современных автоматических устройств. Существующие в
настоящее время линейные и генотипические математические и технические модели
мозга ещё очень упрощённы, имеют слишком большое количество граничных условий,
в них ещё далеко неполно использованы уже известные особенности и
закономерности работы нейронов и нейронных комплексов. Однако и эти модели
представляют ценность для нейрофизиологии, выявляя важные детали, на которые до
сих пор не обращалось внимания.
ЛИТЕРАТУРА
Анохин П.К.
Электроэнцефалографический анализ условного рефлекса. М., Медгиз, 1958.
Беритов И.С. Учение об основных
элементах центральной координации скелетной мускулатуры. Пг., 1916.
Васильев Л.Л. Об основных
функциональных состояниях нервной ткани. В сб: «Новое в рефлексологии и
физиологии нервной системы». 1, 4, Л., 1924.
Введенский Н.Е. Полн. собр. соч.
1–4. Изд. ЛГУ, 1951–1953.
Голиков Н.В. Следовые реакции
нервной системы в зависимости от частоты и силы предшествовавших им
раздражений. Тр. Ленингр. об-ва естествоиспыт, 33, Л., 1957.
Голиков Н.В. Усвоение ритма
раздражений нервными центрами. Раб. физиол. лабор., 11, ЛГУ, 1930.
Голиков Н.В. Функциональные
изменения нерва при локальном стрихнинном отравлении. Тр. Ленингр. об-ва
естествоиспыт., 62, Л., 1933.
Голиков Н.В. Физиологическая
лабильность и её изменения при основных нервных процессах. Изд. ЛГУ, 1950.
Голиков Н.В. Проблема
реактивности организма в свете физиологических исследований. Вестник ЛГУ, 10,
24, 1952.
Голиков Н.В. Физиологические
основы теории электроэнцефалографии. В сб.: «Вопросы теории и практики
электроэнцефалографии», Изд. ЛГУ, 1956.
Голиков Н.В. Роль явлений
синхронизации в образовании системных реакций нервных центров. В сб.: «Проблемы
физиологии и патол. высшей нервной деятельности», 1, Л., Изд. АМН СССР, I960.
Кеннон В. и Розенблют А.
Повышение чувствительности денервированных структур. М., Изд. И. Л., 1951.
Кунстман К.И. и Орбели Л.А. О
последствиях деафферентации задней конечности у собак. «Изв. инст. им.
Лесгафта», 9, 2, 1924.
Ливанов М.Н. Анализ
биоэлектрических процессов в коре головного мозга кролика. «Сов. невропатол. и
психиатрия», 3, 1934.
Ливанов М.Н., Королькова Т.А.,
Френкель Г. М. Электрофизиологические исследования высшей нервной деятельности.
«Журнал высшей нервной деятельности», 1, 1951.
Мнухина Р.С.
Электроэнцефалографическое исследование условнорефлекторных реакций и их анализ
в свете теории Н. Е. Введенского, Изд. ЛГУ, 1964.
Можаева Г.Н. Влияние силы
раздражения на величину местной биоэлектрической реакции нерва. «Биофизика», 3,
1958.
Морелл Ф. Действие поверхностной
анодной поляризации на двигательную реакцию и на характер разрядов отдельных
корковых клеток. «Физиол. журнал СССР», 48, 3, 1962.
Орбели Л.А. Обзор учения о
симпатической иннервации скелетных мышц, органов чувств и центральной нервной
системы. «Физиол. журнал СССР», 15, 1, 1932.
Орбели Л.А. (1937). Лекции по
физиологии нервной системы. Избр. труды, т. II, М.—Л., Изд. АН СССР, 1962.
Орбели Л.А. (1949). Вопросы
высшей нервной деятельности. Избр. труды, т. I, М.—Л., Изд. АН СССР, 1961.
Павлов И.П. (1924). Раздражение и
торможение и экспериментальные неврозы у собак. В сб.: «20-летний опыт изучения
высшей нервной деятельности (поведение животных)», М.—Л., Изд. АН СССР, 1951.
Русинов В.С. Проблема
стационарного возбуждения и изменения постоянного потенциала в коре большого
мозга при доминанте и образовании условного рефлекса. «Журнал высшей нервной деятельности»,
9, 5. М., 1961.
Русинов В.С. О переходе доминанты
в торможение. В сб.: «Рефлексы головного мозга», М., изд. «Наука», 1965.
Сеченов И. М. (1863). Рефлексы
головного мозга. Избр. произв. 1, 1, М.— Л., Изд. АН СССР, 1952.
Сеченов И.М. (1868). Об
электрическом и химическом раздражении чувствующих спинномозговых нервов
лягушки. Избр. произв. 2, М.— Л., Изд. АН СССР, 1956.
Сеченов И.М. (1898).
Физиологические очерки. Избр. труды, 1. Изд. АН СССР, 1952.
Сологуб М.И.
Электрофизиологические показатели лабильности нерва и их изменения при развитии
парабиоза. «Вестник ЛГУ», 13, 97, 1957.
Стёпушкина Т.А. Влияние частичной
деафферентации на функциональное состояние спинномозговых и бульбарных центров.
Вестник ЛГУ, 15, 33, 1962.
Трофимов Л.Г., Любимов Н. Н.,
Наумова Т. С. Взаимодействие коры и подкорки при образовании оборонительных и
пищевых условных рефлексов. Международный коллоквиум по электроэнцефалографии
высш. нерв, деят., М., 1962.
Ухтомский А.А. Собр. соч. 1, 2,
5, Изд. ЛГУ, 1951.
Фессар А. Механизмы межнейронной
синхронизации и их нарушение при эпилептических явлениях. В сб.: «Проблемы
общей нейрофизиологии и высшей нервной. деятельности». Тр. 1 Москов. мед. ин-та
им. И.М. Сеченова, 11, 1961.
Хапажев Т.Ш. Изменения порога для
вызова отрицательных дендритных потенциалов и пороги прямой возбудимости
моторной зоны коры под влиянием стимуляторов и наркотиков. В кн.:
«Электрофизиология нервной системы», Ростов-на-Дону, 1963.
Шевелёва В.С. Эволюция функций
нейронов верхнего шейного симпатического ганглия у кроликов в онтогенезе. 4-е
научн. совещ. по эволюц. физиол. Л., Изд. АН СССР, 1965.
|
|
|
|
|
|
|
|
|